Vermehrung
Bisher wurde kein sicherer Fall geschlechtlicher Fortpflanzung bei Testaceen bekannt. Die Regel ist die ungeschlechtliche Vermehrung durch Zellteilung. Diese wiederholt sich je nach Spezies und Umweltbedingungen in Abstand von ein bis wenigen Tagen und findet meist nachts statt.
Zur Teilung tritt aus der Öffnung der Mutterzelle eine Knospe hervor. Diese Knospe, eine besondere Form eines Pseudopodiums, erreicht bald die Form und Größe der Mutterzelle. (Eigentlich dürfte ich nicht von Mutter und Tochter sprechen. Beide Zellen sind neu. Das Exemplar. welches im alten Gehäuse bleibt, nenne ich Mutter.)
Erst nach Fertigstellung des neuen Gehäuses teilt sich der Zellkern, einer davon wandert in die bereits fertige neue Zelle, und die beiden Einzelzellen trennen sich.
Dagegen wird bekanntlich bei den höherstehenden Eukaryoten die Zellteilung eingeleitet und reguliert durch die Teilung des Zellkerns.

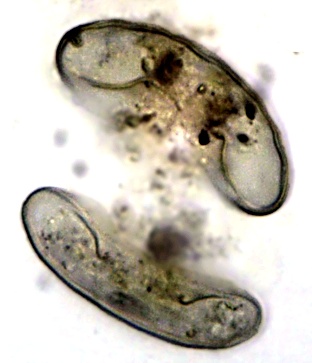
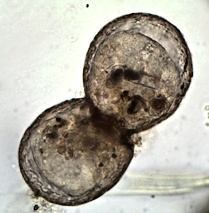
Teilung einer 150 µm hohen Lesquereusia modesta, rechts die dunkle ältere "Mutter"-Schale.
In den Jahren 1916 bis 1918 wurden an der Johns Hopkins University in Baltimore drei aufwändige Studien zur Vererbung äußerer Merkmale bei der ungeschlechtlichen Vermehrung von Difflugia corona, Arcella dentata und Centropyxis aerophila durchgeführt.
Es wurden Modifikationen von Form und Größe bei unzähligen aufeinanderfolgenden Klon-Generationen isolierter Testaceen gemessen und ausgewertet. Da während des Zellteinungsvorgangs der Zellkern noch unbeteiligt blieb, vermutete H. S. Jennings, dass das Zytoplasma und nicht der Zellkern Träger der Vererbung sein könnte:
In this case we should have in Difflugia an organism in which the cytoplasm in place of the nucleus is the 'organ of inheritance;' i.e., is the seat of the diversities that give rise to diversities in the next generation ... during vegetative reproduction“.
und F. M. Root über Centropyxis:
Wilfried Schönborn schrieb 1966:
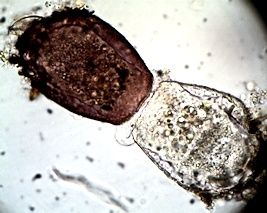
Arcella arenaria (67 µm Durchmesser),
kurz nach der Trennung.
Auch die Verteilung der zur Verstärkung über die Oberfläche verteilten Fremdkörper (Xenosomen) oder selbstgebildeten Plättchen (Idiosomen), ist vor der endgültigen Trennung von Mutter und Tochter abgeschlossen. Zu diesem Zweck hatte die Mutterzelle vorher geeignete Quarzkörner, Diatomeenschalen usw. eingesammelt bzw. kieselige Schuppen selbst gebildet, und diese als Reserve-Bauelemente im Inneren gespeichert. Zur Teilung werden diese Bauelemente in die Knospe transportiert und vor Aushärtung des Pseudochitins äußerst sinnvoll über die Oberfläche verteilt.
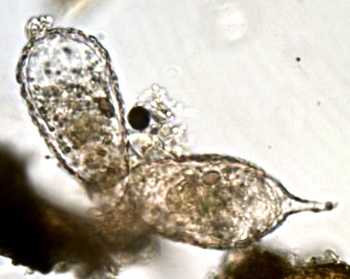
rechtes Exemplar ca. 70 µm mit Spitze.
links vermutlich die Tochter
(Mit kümmerlicher Spitze)
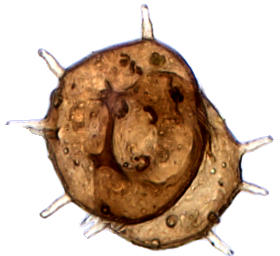
Centropyxis aculeata
Ist das planvolles Vorgehen, tierisches Verhalten, oder ein noch nicht gänzlich geklärter physikalischer Vorgang, bei dem möglicherweise die Oberflächenspannung des aushärtenden Pseudochitins die Partikel verteilt?
Ludwig Rhumbler wies bereits um 1900 nach, dass physikalische Gesetze beteiligt sind, die auch am leblosen Körper wirksam sind. Er impfte Rhizinus/Alkoholtröpfchen (im Wasser) mit winzigen Steinchen. Diese verteilten sich umgehend durch die Oberflächenspannung gleichmäßig über die Oberfläche der Tröpfchen, wie das auch bei der Schalenbildung der Difflugien geschieht.
Damit ist allerdings nicht erklärt, wie als Vorbereitung der Teilung eine „intelligente“ Auswahl der Baumaterialien stattfinden konnte, und wie diese sich je nach Form und Größe gezielt an verschiedenen Stellen der neuen Schale platzieren.
Nach der Teilung ändern sich die Merkmale der Individuen nicht mehr, die neue Zelle im neuen Gehäuse ist fertig. Lediglich das zunächst hell durchscheinende Pseudochitin verfärbt sich im Laufe der Alterung durch Metalleinlagerungen bräunlich-rötlich und wird undurchsichtig.
Die zeitlichen Abläufe: (unter günstigen Bedingungen)
Die Bildung einer neuen Schale vom Austritt der Knospe bis zur Armierung des fertigen Gehäuses dauert etwa 30 bis 90 Minuten .
Danach zirkuliert das durch Wasseraufnahme vergrößerte Plasma lebhaft zwischen den beiden Gehäusen. Der Zellkern teilt sich, und einer wandert in die neue Schale. Danach trennen sich die beiden Zellen. Dauer des Gesamtvorgangs ca. 2 Stunden.
Verdoppelung der Population etwa alle 2-3 Tage.
Diese Zeitangaben variieren erheblich je nach Spezies und Umweltbedingungen und sollen hier nur als Anhaltspunkt dienen.
-------------------
(Quellen: Schönborn 1966 und weitere am Schluss)