Systematik
Die Testaceen (deutsch Schalenamöben) zählt man neben den Nacktamöben, Schleimpilzen, Foraminiferen und Strahlentierchen zu den Amöbozoa: Mikroskopisch kleine, sehr einfach gebaute eukaryotische Einzeller mit besonderen gemeinsamen Eigenschaften. Amoebozoa bilden eine Lebensform oder Organisationsstufe, aber keine taxonomische Gruppe.
Die phylogenetische Position der Testaceen ist anhand der DNA-Sequenzen gesichert. Arcellinida und Euglyphida sind jedoch genetisch weit voneinander entfernte Gruppen. Die ältesten Testaceen-Gehäuse wurden für das Erdzeitalter Cryogenium (720 - 635 Mya, also vor der Kambrischen Explosion und noch vor dem Ediacarium) nachgewiesen.
Hoogenrad u. De Groot (1935):
"viele Rhizopodenarten zeigen eine so große
Variabilität und eine so starke Aufsplitterung [ ... ] dass manche neuaufgestellte Arten nur ganz willkürliche Ausschnitte aus einer sehr großen Formenreihe darstellen".
Ralf Meisterfeld formulierte 1979:
"Die Abgrenzung des Artbegriffs bei den sich ungeschlechtlich fortpflanzenden Thekamoben stößt auf grundsätzliche Schwierigkeiten. Die Individuen einer Art gehören verschiedenen Linien an, deren Genbestand isoliert ist. Ein genetisch bestimmter
Artbegriff ist also bei den Testaceen nicht anwendbar. Im Laufe der Evolution entfernen sich die einzelnen Linien voneinander, was sich zunächst in kleinen morphologischen Unterschieden bemerkbar macht. Den Zeitpunkt, zu dem sich eine solche Linie so weit von einer anderen entfernt hat, daß man von einer Unterart oder Art sprechen kann, haben die einzelnen Spezialisten sehr unterschiedlich festgesetzt."
Mehr Klarheit versprechen erst die laufenden genetischen Vergleiche.
Lumper oder Splitter
„Diese beiden Wörter (Splitter = 'Spalter' und Lumper = 'In-einen-Topf-Werfer') sind in der biologischen Systematik zu Fachbegriffen geworden: Ein Lumper fasst Tiere (oder Pflanzen) zu großen Gruppen zusammen, ein Splitter teilt sie in viele kleine Gruppen und Arten auf.
Splitter erfinden immer neue Namen und erheben in Extremfällen ... fast jeden neu entdeckten Fund in den Rang einer eigenen Spezies“.
Richard Dawkins
Unterordnung Glutinoconcha (Lahr et al 2019)
- Sphaerothecina (rund kreissymmetrisch, teller- bis halbkugelförmig, zentraler Mund)
- Arcellidae
- Netzelidae
- Trigonopyxis
- und andere
- Longithecina
(länglich, Durchschnitt und endständiger Mund rund, bedeckt mit Fremdmaterial oder selbsterzeugten Partikeln)
- Difflugiidae
- Lesquereusiidae
- Excentrostoma (Dorso-ventral abgeflacht, Mund exzentrisch)
- Centropyxidae
- Plagiopyxidae
- Hyalospheniformis (flach, birnen- oder vasenförmig, Schale unbedeckt oder mit erbeuteten Xenosomen, Mund endständig)
- Hyalosphenia
- Nebela
- Planocarina
- Quadrulella
- Mrabella
- Gibbocarina
- Longinebela
- und andere
- Volnustoma Flach, meist mit Xenosomen, mit endständiger Schlitzöffnung.
- Heleoperidae
Unterordnung Organoconcha (Lahr et al 2019)
- Microclamidae
- Microclamys
- Pyxidicula
Unterordnung Phryganellina (Bovee 1985)
- Phryganellidae
- Phryganella
- Cryprodifflugidae
- Cryptodifflugia
- Wailesella
Die filosen Testaceen (nach Heger et.al. 2010):
Ordnung Euglyphida
- Euglyphina
- Assulinidae
- Sphenoderiidae
- Trimatidae
- Euglyphydae
- Tracheloglypha
- Cyphoderiidae
- Paulinellidae
Ordnung Amphitremida
(mit zwei gegenständigen Pseudostomen.Mein Artikel über Torfmoos bewohnende Testaceen im Micscape eZine.
Zoochlorellen
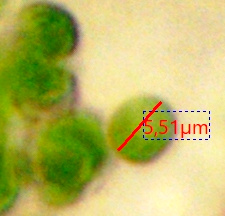
Mehrere Testaceen-Gattungen in nährstoffarmen Gewässern ernähren sich mixotrophisch.
Diese Arten, beispielsweise Archerella flavum, Hyalosphenia papilio, Heleopera sphagni, Amphitrema wrigthianum, Cochliopodium, einige große Difflugien, Arcella artocrea (bei Leidy), Placocista spinosa (Gomaa et al), die Nacktamöbe Mayorella viridis, können Nahrung durch Fressen anderer Organismen aufnehmen, sich aber auch durch symbiontische Algen per Fotosynthese versorgen lassen (Mixotrophie). Bei der Symbiose handelt es sich vornehmlich um den Austausch von Assimilationsprodukten; die Algen empfangen vom Wirt Kohlendioxid und Aminosäuren, während sie im Gegenzug Zucker aus ihrer Photosynthese abgeben. Es handelt sich auch um eine "Hungerversicherung" für den Wirt, die es ihm ermöglicht, Perioden mit geringer Nahrungsverfügbarkeit zu überstehen.
Diese Wirte besitzen in der Regel ein sehr durchscheinendes Gehäuse ohne Xenosomen und lassen so ungehindert Licht an ihre Symbionten. Ausnahme bilden hier die genannten großen Difflugien, deren Schalen mit Xenosomen besetzt und so fast undurchsichtig sind.
Die Algen bleiben auch im Zusammenleben mit ihrem Wirt selbständige Organismen, die allein lebensfähig sind. Die Wirte können dagegen in der Regel nicht ohne diese Symbionten überleben. Offensichtlich wird bei der Zellteilung der Algenvorrat auf die beiden entstehenden Individuen aufgeteilt.
Die Symbionten werden sowohl bei der Zellteilung auf beide Töchter aufgeteilt und vermehren sich dort weiter. Aber sie besiedeln anscheinend auch aus einem freischwimmenden Stadium geeignete Testaceen. Wie sie es anstellen, nicht verdaut zu werden, ist rätselhaft. In Notzeiten greifen z.B Archerella flavum wohl doch auch auf diesen Vorrat zu und verdauen sie teilweise.
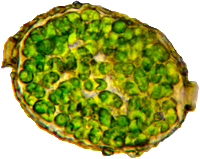
Beispiel Amphitrema wrightianum.
Diese Symbionten sind alles nahe miteinander verwandte, 5-6 µm (meine Messungen) große Algen aus der Gattung Chlorella, wie sie auch in der Nacktamöbe Mayorella, im grünen Pantoffeltier, in der Süßwasserhydra und in Flechten leben = TACS (Testate Amoeba Chlorella Symbionts).
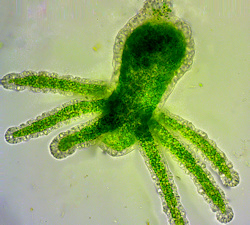
Beispiel Grüne Hydra.
---------------
Die einzige bekannte Ausnahme bildet die ca. 30 µm große Euglyphide Paulinella chromatophora. Jede Paulinella beherbergt zwei so genannte Chromatophoren, einstige Cyanobakterien. Diese haben Teile ihres Genoms an den Zellkern des Wirts transferiert und sind somit zu Organellen geworden. Jede Paulinella beherbergt zwei dieser Endosymbionten. Bei der Teilung erhält jede Teilzelle eine der beiden Organellen, die sich umgehend durch Verdoppelung komplettieren. P. chromatophora leben ausschliesslich von den Fotosyntheseprodukten der Symbionten. Diese Symbiose ist erdgeschichtlich jung und fand vor etwa 60 Millionen Jahren statt.
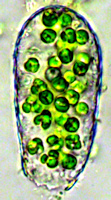
Hier haben sich (ebenfalls etwa 5 - 6 µm große) Chlorella-Algen in einer leeren Euglypha-Schale eingerichtet. Haben diese Organismen aktiv den Schutz der leeren, hyalinen Testaceenschale gesucht? Ist hier möglicherweise der Ursprung der Symbiose der Algen mit Schalenamöben bzw. den oben genannten anderen Ein- oder Wenigzellern zu vermuten?